



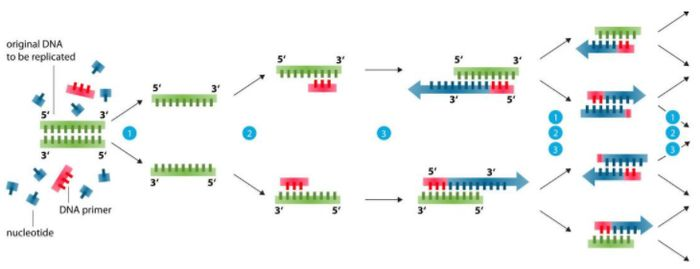
Základ návrhu základního nátěru (99 % problémů lze vyřešit)
1. Délka primeru: Učebnice vyžaduje 15-30bp, obvykle asi 20bp.Skutečná podmínka je lepší být 18-24 bp, aby byla zajištěna specificita, ale čím delší, tím lepší, příliš dlouhý primer také sníží specificitu a sníží výtěžek.
2. Rozsah amplifikace primeru: Vhodné je 200-500 bp a fragment lze za specifických podmínek rozšířit na 10 kb.
3. Základní báze: Obsah G+C by měl být 40-60 %, příliš malý efekt zesílení G+C není dobrý, příliš mnoho G+C snadno vytvoří nespecifické pásy.ATGC je nejlépe distribuován náhodně, vyhýbá se shlukům více než 5 purinových nebo pyrimidinových nukleotidů.Multi-gc pro 5′ konec a mezilehlé sekvence pro zvýšení stability, vyhněte se bohaté GC na 3′ konci, žádné GC na posledních 3 bázích nebo žádné GC na 3 z posledních 5 bází.
4. Vyhněte se sekundární struktuře v primerech a vyhněte se komplementaci mezi dvěma primery, zejména komplementaci na 3' konci, jinak se vytvoří dimer primeru a vytvoří se nespecifické amplifikované pásy.
5. Báze na 3' konci primerů, zejména poslední a předposlední báze, by měly být přísně spárovány, aby se zabránilo selhání PCR kvůli nespárovaným koncovým bázím.
6. Primery mají nebo mohou být přidány s vhodnými místy štěpení a amplifikovaná cílová sekvence by výhodně měla mít vhodná místa štěpení, což je velmi výhodné pro analýzu štěpení nebo molekulární klonování.
7. Specifičnost primerů: primery by neměly mít žádnou zjevnou homologii s jinými sekvencemi v databázi sekvencí nukleových kyselin.
8. Naučte se používat software: PP5, Oligo6, DNAstar, Vector NTI, primer3 (Tento online design funguje nejlépe).
Výše uvedený obsah může vyřešit alespoň 99 % problémů s návrhem primeru.
Kontrolujte detaily návrhu primeru
1. Délka primeru
Obecná délka primeru je 18~30 bází.Obecně je nejdůležitějším faktorem určujícím teplotu nasedání primeru délka primeru.Teplota žíhání primeru je obecně zvolena (hodnota Tm -5 °C) a některé přímo používají hodnotu Tm.Pro přibližný výpočet teploty žíhání primerů lze použít následující vzorce.
Když je délka primeru menší než 20 bp: [4(G+C)+2(A+T)]-5℃
Když je délka primeru větší než 20 bp: 62,3℃+0,41℃(%GC)-500/délka-5℃
Kromě toho lze pro výpočet teploty žíhání použít také mnoho software, princip výpočtu bude jiný, takže někdy může mít vypočítaná hodnota malou mezeru.Pro optimalizaci PCR reakcí se pro nejlepší účinnost a specificitu používají nejkratší primery, které zajišťují teploty nasedání ne nižší než 54 °C.
Celkově vzrůstá specificita primeru čtyřnásobně pro každý další nukleotid, takže minimální délka primeru pro většinu aplikací je 18 nukleotidů.Horní hranice délky primeru není příliš důležitá, hlavně souvisí s účinností reakce.Vzhledem k entropii platí, že čím delší je primer, tím nižší je rychlost, jakou nasedá, aby se navázal na cílovou DNA a vytvořil stabilní dvouvláknový templát pro vazbu DNA polymerázy.
Při použití softwaru pro návrh primerů lze délku primerů určit podle hodnoty TM, zejména u primerů fluorescenční kvantitativní PCR by měla být kontrolována TM=60 °C nebo podobně.
2. Obsah GC
Obecně je obsah G+C v sekvencích primerů 40 %~60 % a obsah GC a hodnota Tm páru primerů by měly být koordinovány.Pokud má primer závažnou tendenci ke GC nebo AT, lze na 5' konec primeru přidat příslušné množství A, T nebo G a C konce.
3. Teplota žíhání
Teplota žíhání by měla být o 5 ℃ nižší než teplota odřetězení.Pokud je počet bází primerů malý, lze teplotu nasedání vhodně zvýšit, což může zvýšit specificitu PCR.Pokud je počet bází velký, lze teplotu žíhání přiměřeně snížit.Rozdíl teplot nasedání mezi párem primerů 4 °C ~ 6 °C neovlivní výtěžek PCR, ale v ideálním případě je teplota nasedání páru primerů stejná, která se může pohybovat mezi 55 °C ~ 75 °C.
4. Vyhněte se oblasti sekundární struktury amplifikační šablony
Při výběru amplifikovaného fragmentu je nejlepší vyhnout se oblasti sekundární struktury templátu.Stabilní sekundární struktura cílového fragmentu může být predikována a odhadnuta příslušným počítačovým softwarem, což je užitečné pro výběr templátu.Experimentální výsledky ukazují, že expanze je často neúspěšná, když je volná energie (△G) oblasti, která se má expandovat, menší než 58,6 kJ/mol.
5. Neshoda s cílovou DNA
Když je amplifikovaná cílová DNA sekvence velká, primer se může vázat na více částí cílové DNA, což vede k tomu, že se ve výsledku objeví více pásů.Tentokrát je nutné využít testování softwaru BLAST, web:http://www.ncbi.nlm.nih.gov/BLAST/.Vyberte Zarovnat dvě sekvence (bl2seq).
Vkládání sekvencí primerů do zóny 1 a cílových sekvencí DNA do zóny 2 je vzájemně zaměnitelné a BLAST vypočítává komplementární, antisense a další možnosti, takže uživatelé si nemusí všímat, zda jsou oba řetězce sense řetězce.Můžete také zadat číslo GI, pokud znáte číslo GI sekvence v databázi, takže nemusíte vkládat velkou část sekvence.Nakonec klikněte na Zarovnat na 3, abyste viděli, zda má primer více homologních míst v cílové DNA.
6. Terminál primeru
3' konec primeru je místo, kde začíná rozšíření, takže je důležité zabránit tomu, aby tam začaly neshody.3' konec by neměl být více než 3 po sobě jdoucí G nebo C, protože to způsobí, že primer bude chybně spuštěn v oblasti sekvence obohacené G+C.3' konec nemůže tvořit žádnou sekundární strukturu, s výjimkou speciálních reakcí PCR (AS-PCR), kdy 3' konec primeru nemůže být špatně spárován.Například, pokud je kódující oblast amplifikována, 3'konec primeru by neměl být ukončen na třetí pozici kodonu, protože třetí pozice kodonu je náchylná k degeneraci, což ovlivní specificitu a účinnost amplifikace.Při použití anexových primerů se podívejte do tabulky použití kodonů, věnujte pozornost biologické preferenci, nepoužívejte anexní primery na 3′ konci a použijte vyšší koncentraci primerů (1uM-3uM).
7. Sekundární struktura primerů
Samotné primery by neměly mít komplementární sekvence, jinak se primery samotné složí do vlásenkových struktur a tato sekundární struktura ovlivní vazbu primerů a templátů v důsledku sterické zábrany.Pokud se použije umělé posouzení, kontinuální komplementární báze samotných primerů by neměly být větší než 3 bp.Mezi těmito dvěma primery by neměla být žádná komplementarita, zejména by se mělo zabránit komplementárnímu překrývání 3' konce, aby se zabránilo tvorbě dimerů primerů.Obecně by mezi párem primerů nemělo existovat více než 4 po sobě jdoucí báze homologie nebo komplementarita.
8. Přidejte značky nebo lokusy
5' konec má malý vliv na amplifikační specificitu, a proto může být modifikován bez ovlivnění amplifikační specifity.Modifikace primeru 5' konec zahrnovala: přidání enzymového restrikčního místa;Značený biotin, fluorescence, digoxin, Eu3+ atd. Zavést sekvence DNA vázající protein;Zavedení mutačních míst, vložení a chybějící mutační sekvence a zavedení promotorových sekvencí atd. Extra báze více či méně ovlivní účinnost amplifikace a zvýší šanci na tvorbu primeru dimeru, ale pro další krok je třeba udělat určité ústupky.Další sekvence, které neexistují na cílové sekvenci, jako jsou restrikční místa a promotorové sekvence, mohou být přidány na 5' konec primeru bez ovlivnění specificity.Tyto sekvence nejsou zahrnuty do výpočtu hodnot Tm primeru, ale měly by být testovány na komplementaritu a vnitřní sekundární strukturu.
9. Subklony
Většinu času je PCR pouze předběžné klonování a pak potřebujeme subklonovat cílový fragment do různých vektorů, takže musíme navrhnout další báze pro další operaci v kroku PCR.
Některé sekvence navržené pro subklonování jsou shrnuty níže.
Bylo přidáno restrikční místo pro restrikční endonukleázu
Přidání enzymových restrikčních míst je nejběžněji používanou metodou pro subklonování produktů PCR.Obecně je místo štěpení šest bází, kromě 5' konce místa štěpení je třeba přidat 2 až 3 ochranné báze.Počet ochranných bází požadovaných různými enzymy je však různý.Například SalⅠ nevyžaduje žádnou ochrannou základnu, EcoRⅤ vyžaduje 1 ochrannou základnu, NotⅠ vyžaduje 2 ochranné základny a Hind Ⅲ vyžaduje 3 ochranné základny.
LIC přidává ocas
Úplný název LIC je klonování nezávislé na ligaci, metoda klonování vynalezená společností Navogen specificky pro její část vektoru pET.Nosič pET připravený metodou LIC má nekomplementární jednořetězcové lepivé konce o 12-15 bázích, které doplňují odpovídající lepivé konce na fragmentu cílového inzertu.Pro účely amplifikace by sekvence primeru 5' vloženého fragmentu měla doplňovat vektor LIC.3′→5′ extranektní aktivita T4 DNA polymerázy může po krátké době vytvořit na vloženém fragmentu jednovláknový lepivý konec.Protože produkt lze vytvořit pouze vzájemným nasedáním připraveného inzertního fragmentu a vektoru, je tato metoda velmi rychlá a účinná a jedná se o přímé klonování.
Řízený klon TA přidat ocas
TA klonování nebylo schopno zacílit fragment do vektoru, takže později Invitrogen zavedl vektor, který mohl cílit klonování, který obsahoval čtyři prominentní báze GTGGS na jednom konci.Proto by při návrhu PCR primerů měly být odpovídajícím způsobem přidány komplementární sekvence, aby bylo možné fragmenty „orientovat“.
Pokud máte málo času, můžete zkusit přímou syntézu, kombinování genu s vektorem, což u musekularistů nazýváme syntézou ET genu.
D. Metoda in-fúzního klonování
Není nutná ligáza, není nutná dlouhá reakce.Dokud je v návrhu primerů zavedena sekvence na obou koncích linearizovaného vektoru, pak se produkt PCR a linearizovaný vektor přidají do infúzního enzymového roztoku obsahujícího BSA a umístí se na půl hodiny při pokojové teplotě, lze provést transformaci.Tato metoda je zvláště vhodná pro konverzi velkých objemů.
10. Sloučit primer
Někdy jsou o návrhu primeru známy pouze omezené informace o sekvenci.Například, pokud je známa pouze aminokyselinová sekvence, může být navržen slučovací primer.Slučovací primer je směs různých sekvencí představujících všechny různé možnosti bází, které kódují jednu aminokyselinu.Chcete-li zvýšit specifitu, můžete se obrátit na tabulku použití kodonů, abyste snížili připojení podle preferencí použití báze různých organismů.Hypoxanthin může být spárován se všemi bázemi, aby se snížila teplota nasedání primeru.Nepoužívejte připojené báze na 3′ konci primeru, protože nasedání posledních 3 bází na 3′ konci je dostatečné k zahájení PCR na nesprávném místě.Používají se vyšší koncentrace primerů (1μM až 3μM), protože primery v mnoha anexi směsích nejsou specifické pro cílový templát.
Suroviny pro PCRřízení
1. Množství primeru
Koncentrace každého primeru je 0,1 ~ 1 umol nebo 10 ~ 100 pmol.Je lepší dosáhnout požadovaného výsledku s co nejmenším množstvím primeru.Vysoká koncentrace primeru způsobí nesprávné párování a nespecifickou amplifikaci a zvýší možnost tvorby dimerů mezi primery.
2. Koncentrace primeru
Koncentrace primerů ovlivňuje specificitu.Optimální koncentrace primeru je obecně mezi 0,1 a 0,5 μM.Vyšší koncentrace primerů vedou k amplifikaci nespecifických produktů.
3. Teplota žíhání základního nátěru
Dalším důležitým parametrem pro primery je teplota tání (Tm).Toto je teplota, kdy je 50 % primerů a komplementárních sekvencí reprezentováno jako dvouvláknové molekuly DNA.K nastavení teploty nasedání PCR je vyžadována Tm.V ideálním případě je teplota nasedání dostatečně nízká k zajištění účinného nasedání primerů s cílovou sekvencí, ale dostatečně vysoká, aby se snížila nespecifická vazba.Přiměřená teplota žíhání od 55 ℃ do 70 ℃.Teplota žíhání je obecně nastavena o 5 °C nižší než Tm primeru.
Existuje několik vzorců pro nastavení Tm, které se značně liší v závislosti na použitém vzorci a sekvenci primerů.Protože většina vzorců poskytuje odhadovanou hodnotu Tm, jsou všechny teploty žíhání pouze výchozím bodem.Specificitu lze zlepšit analýzou několika reakcí, které postupně zvyšují teplotu žíhání.Začněte pod odhadovanou Tm-5℃ a postupně zvyšujte teplotu žíhání v krocích po 2℃.Vyšší teplota žíhání sníží tvorbu primerových dimerů a nespecifických produktů.Pro nejlepší výsledky by měly mít dva primery přibližné hodnoty Tm.Pokud je rozdíl Tm párů primerů větší než 5 °C, primery budou vykazovat významný falešný start při použití nižší teploty nasedání v cyklu.Pokud se dva primery Tm liší, nastavte teplotu žíhání o 5 °C nižší, než je nejnižší Tm.Alternativně, pro zvýšení specifity, může být nejprve provedeno pět cyklů při teplotách nasedání navržených pro vyšší Tm, následované zbývajícími cykly při teplotách nasedání navržených pro nižší Tm.To umožňuje získat částečnou kopii cílové šablony za ztížených podmínek.
4. Čistota a stabilita primeru
Standardní čistota zakázkových primerů je dostatečná pro většinu aplikací PCR.Odstranění benzoylových a isobutylylových skupin odsolením je minimální, a proto neinterferuje s PCR.Některé aplikace vyžadují purifikaci k odstranění všech sekvencí, které nemají plnou délku v procesu syntézy.Tyto zkrácené sekvence se vyskytují, protože účinnost chemie syntézy DNA není 100 %.Jedná se o kruhový proces, který využívá opakované chemické reakce, protože každá báze se přidává, aby se vytvořila DNA od 3′ do 5′.V obou cyklech můžete selhat.Delší primery, zejména ty větší než 50 bází, mají velký podíl zkrácených sekvencí a mohou vyžadovat purifikaci.
Výtěžek primerů je ovlivněn účinností syntetické chemie a metodou čištění.Biofarmaceutické společnosti, jako je Cytology a Shengong, všechny používají minimální jednotku OD k zajištění celkové produkce oligonukleosidů.Vlastní základní nátěry jsou dodávány ve formě suchého prášku.Primery je nejlepší znovu rozpustit v TE tak, aby výsledná koncentrace byla 100μM.TE je lepší než deionizovaná voda, protože pH vody je často kyselé a způsobí hydrolýzu oligonukleosidů.
Stabilita primerů závisí na podmínkách skladování.Suchý prášek a rozpuštěné primery by měly být skladovány při -20 °C.Primery rozpuštěné v TE v koncentracích vyšších než 10μM mohou být stabilně skladovány při -20 °C po dobu 6 měsíců, ale mohou být skladovány pouze při pokojové teplotě (15 °C až 30 °C) po dobu kratší než 1 týden.Suché práškové základní nátěry lze skladovat při -20 C po dobu nejméně 1 roku a při pokojové teplotě (15 C až 30 C) až 2 měsíce.
5. Enzymy a jejich koncentrace
V současnosti je Taq DNA polymeráza v podstatě enzym genového inženýrství syntetizovaný koliformními bakteriemi.Množství enzymu potřebného ke katalýze typické PCR reakce je přibližně 2,5 U (vztahuje se na celkový reakční objem 100 ul).Pokud je koncentrace příliš vysoká, může to vést k nespecifické amplifikaci;pokud je koncentrace příliš nízká, sníží se množství syntetického produktu.
6. Kvalita a koncentrace dNTP
Kvalita dNTP úzce souvisí s koncentrací a účinností PCR amplifikace.Prášek dNTP je zrnitý a jeho variabilita ztrácí svou biologickou aktivitu, pokud je nesprávně skladován.Roztok dNTP je kyselý a měl by být používán ve vysoké koncentraci, s 1M NaOH nebo 1M pufrovým roztokem Tris.HCL pro úpravu jeho pH na 7,0 ~ 7,5, malé množství dílčího balení, skladování ve zmrazeném stavu při -20 °C.Vícenásobné zmrazení-rozmrazení degraduje dNTP.V PCR reakci by měl být dNTP 50 ~ 200 umol/l.Zejména je třeba věnovat pozornost tomu, aby koncentrace čtyř DNTPS byla stejná (stejná molární příprava).Pokud se koncentrace kteréhokoli z nich liší od ostatních (vyšší nebo nižší), dojde k nesouladu.Příliš nízká koncentrace sníží výtěžek produktů PCR.dNTP se může kombinovat s Mg2+ a snižovat koncentraci volného Mg2+.
7. Templátová (cílový gen) nukleová kyselina
Množství a stupeň purifikace templátové nukleové kyseliny je jedním z klíčových vazeb pro úspěch nebo neúspěch PCR.Tradiční metody čištění DNA obvykle používají SDS a proteázu K k trávení a likvidaci vzorků.Hlavní funkce SDS jsou: rozpouštět lipidy a proteiny na buněčné membráně, tím ničit buněčnou membránu rozpuštěním membránových proteinů a disociovat jaderné proteiny v buňce, SDS se může také spojovat s proteiny a vysrážet se;Proteáza K může hydrolyzovat a trávit proteiny, zejména histony vázané s DNA, a poté použít organické rozpouštědlo fenol a chloroform k extrakci proteinů a dalších buněčných složek a použít ethanol nebo isopropylalkohol k vysrážení nukleové kyseliny.Extrahovaná nukleová kyselina může být použita jako templát pro PCR reakce.Pro obecné klinické detekční vzorky lze použít rychlou a jednoduchou metodu k rozpuštění buněk, lyzaci patogenů, štěpení a odstranění proteinů z chromozomů k uvolnění cílových genů a přímo k použití pro PCR amplifikaci.Extrakce templátu RNA obvykle používá guanidinisothiokyanát nebo metodu proteázy K, aby se zabránilo RNáze degradovat RNA.
8. Koncentrace Mg2+
Mg2+ má významný vliv na specificitu a výtěžnost PCR amplifikace.V obecné PCR reakci, když je koncentrace různých dNTP 200 umol/l, vhodná koncentrace Mg2+ je 1,5 ~ 2,0 mmol/l.Koncentrace Mg2+ je příliš vysoká, snižuje se specifita reakce, dochází k nespecifické amplifikaci, příliš nízká koncentrace snižuje aktivitu Taq DNA polymerázy, což má za následek redukci reakčních produktů.
Ionty hořčíku ovlivňují několik aspektů PCR, jako je aktivita DNA polymerázy, která ovlivňuje výtěžek;Dalším příkladem je nasedání primerů, které ovlivňuje specificitu.dNTP a templát se vážou na ionty hořčíku, čímž se snižuje množství volného iontu hořčíku potřebného pro aktivitu enzymu.Optimální koncentrace hořčíkových iontů se liší pro různé páry primerů a templáty, ale typická výchozí koncentrace pro PCR s 200 μM dNTP je 1,5 mM (poznámka: Pro kvantitativní PCR v reálném čase použijte 3 až 5 mM roztok hořčíkových iontů s fluorescenční sondou).Vyšší koncentrace volných iontů hořčíku zvyšují výtěžek, ale také zvyšují nespecifickou amplifikaci a snižují věrnost.Pro stanovení optimální koncentrace byly titrace hořčíkových iontů prováděny v přírůstcích 0,5 mM od 1 mM do 3 mM.Pro snížení závislosti na optimalizaci hořčíkových iontů lze použít Platinum Taq DNA polymerázu.Platinová Taq DNA polymeráza je schopna zachovat funkci v širším rozsahu koncentrací hořčíkových iontů než Taq DNA polymeráza, a proto vyžaduje méně optimalizace.
9. Přísady podporující Pcr
Optimalizace teploty žíhání, návrhu primeru a koncentrace hořčíkových iontů je dostatečná pro vysoce specifickou amplifikaci většiny templátů;některé šablony, včetně šablon s vysokým obsahem GC, však vyžadují další opatření.Aditiva, která ovlivňují teplotu tání DNA, poskytují další způsob, jak zlepšit specifitu produktu a výtěžek.Pro dosažení nejlepších výsledků je nutná úplná denaturace šablony.
Kromě toho sekundární struktura zabraňuje navázání primeru a extenzi enzymu.
PCR aditiva, včetně formamidu, DMSO, glycerinu, betainu a PCRx Enhancer Solution, zvyšují amplifikaci.Jejich možným mechanismem je snížení teploty tání, čímž napomáhá nasedání primerů a napomáhá extenzi DNA polymerázy přes oblast sekundární struktury.PCRx Solution má další výhody.Při použití s DNA polymerázou Platinum Taq a DNA polymerázou Platinum Pfx je vyžadována minimální optimalizace hořčíkových iontů.Technika Platinum je tedy kombinována s aditivem pro zvýšení specificity a zároveň snížení závislosti na třetím přístupu, optimalizaci hořčíkových iontů.Pro dosažení nejlepších výsledků by měla být optimalizována koncentrace aditiv, zejména DMSO, formamidu a glycerolu, které inhibují Taq DNA polymerázu.

10. Horký start
Hot start PCR je jednou z nejdůležitějších metod pro zlepšení specifity PCR vedle dobrého návrhu primeru.Ačkoli optimální teplota prodlužování Taq DNA polymerázy je 72 °C, polymeráza zůstává aktivní při pokojové teplotě.Nespecifické produkty tedy vznikají, když je udržovací teplota nižší než teplota žíhání během přípravy PCR reakce a na začátku tepelného cyklu.Jakmile jsou tyto nespecifické produkty vytvořeny, účinně se amplifikují.Hot-start PCR je zvláště účinná, když jsou místa použitá pro návrh primerů omezena umístěním genetických elementů, jako jsou místně cílené mutace, expresní klonování nebo konstrukce a manipulace genetických elementů používaných pro DNA inženýrství.
Běžnou metodou k omezení aktivity Taq DNA polymerázy je připravit reakční roztok pro PCR na ledu a umístit jej do předehřátého PCR přístroje.Tato metoda je jednoduchá a nenákladná, ale nedokončuje aktivitu enzymu, a proto zcela nevylučuje amplifikaci nespecifických produktů.
Tepelná aktivace zpožďuje syntézu DNA inhibicí základní složky, dokud PCR aparát nedosáhne denaturační teploty.Většina manuálních metod tepelné iniciace, včetně zpožděného přidání Taq DNA polymerázy, je těžkopádná, zejména pro vysoce výkonné aplikace.Jiné metody tepelného primárního nátěru používají voskový štít k uzavření základní složky, včetně hořčíkových iontů nebo enzymů, nebo k fyzické izolaci reaktivních složek, jako jsou templáty a pufry.Během tepelného cyklu se různé složky uvolňují a mísí se dohromady, jak se vosk roztaví.Stejně jako ruční metoda horkého startu je metoda voskového štítu těžkopádná a náchylná ke kontaminaci a není vhodná pro vysoce výkonné aplikace.
Platinová DNA polymeráza je vhodná a účinná pro automatické hot start PCR.Platinum Taq DNA polymeráza se skládá z rekombinantní Taq DNA polymerázy kombinované s monoklonální protilátkou proti Taq DNA polymeráze.Protilátky jsou formulovány pomocí PCR, aby inhibovaly aktivitu enzymu během dlouhodobého udržování teploty.Taq DNA polymeráza byla uvolněna do reakce během 94 °C izolace v denaturačním kroku, čímž byla obnovena plná polymerázová aktivita.Na rozdíl od chemicky modifikované Taq DNA polymerázy pro tepelnou iniciaci, Platinum enzym nevyžaduje k aktivaci polymerázy prodlouženou izolaci při 94 °C (10 až 15 minut).S PlatinumTaq DNA polymerázou bylo 90 % aktivity Taq DNA polymerázy obnoveno po 2 minutách při 94 °C.

11. Nest-PCR
Po sobě jdoucí kola amplifikace pomocí vnořených primerů mohou zlepšit specificitu a citlivost.První kolo je standardní zesílení 15 až 20 cyklů.Malá frakce počátečního amplifikačního produktu byla zředěna 100 až 1000 krát a přidána do druhého kola amplifikace na 15 až 20 cyklů.Alternativně lze velikost počátečního amplifikovaných produktů upravit gelovým čištěním.Ve druhém kole amplifikace se používá nested primer, který se může vázat na cílovou sekvenci uvnitř prvního primeru.Použití nested PCR snižuje možnost amplifikace více cílových míst, protože existuje málo cílových sekvencí komplementárních k oběma sadám primerů.Stejný celkový počet cyklů (30 až 40) se stejnými primery amplifikoval nespecifická místa.Nested PCR zvyšuje citlivost omezených cílových sekvencí (např. vzácné mrnas) a zlepšuje specificitu obtížných PCRS (např. 5′ RACE).
12. Sestupná PCR
Sestupná PCR zlepšuje specificitu použitím podmínek těsného nasedání pro prvních několik cyklů PCR.Cyklus začíná při teplotě žíhání přibližně o 5 °C vyšší, než je odhadovaná Tm, pak se každý cyklus snižuje o 1 °C až 2 °C, dokud teplota žíhání neklesne pod Tm 5 °C.Amplifikovaný bude pouze cílový templát s nejvyšší homologií.Tyto produkty pokračují v expanzi v následujících cyklech a vytlačují zesílené nespecifické produkty.Descending PCR je užitečná pro metody, kde není znám stupeň homologie mezi primerem a cílovým templátem, jako je AFLP DNA fingerprinting.
Související sady PCR
2× hrdina PCRTMMix systém má vyšší toleranci k inhibitorům PCR než běžný systém PCR Mix a snadno si poradí s PCR amplifikací různých komplexních templátů.Díky jedinečnému reakčnímu systému a vysoce účinnému Taq Hero má PCR reakce vyšší účinnost amplifikace, specificitu a citlivost.
Vyšší účinnost zesílení
Má 5'→3' DNA polymerázovou aktivitu a 5'→3' exonukleázovou aktivitu, bez 3'→5' exonukleázové aktivity.
Real Time PCR Easyᵀᴹ-SYBR Green I Kit
Specifický – optimalizovaný pufr a enzym Taq s horkým startem mohou zabránit nespecifické amplifikaci a tvorbě dimeru primeru
Vysoká citlivost – dokáže detekovat nízké kopie šablony
Souprava využívá unikátní reverzní transkripční činidlo Foregene a Foregene HotStar Taq DNA polymerázu v kombinaci s unikátním reakčním systémem pro účinné zlepšení účinnosti amplifikace a specifičnosti reakce.
Čas odeslání: květen-09-2023



